项目文章IF:14.9 | 香蕉 “癌症” 有了绿色新解法:海洋链霉菌靠信号分子,召集有益菌守住根部防线自动草稿

项目文章 | 氮肥过量竟会劣变稻米品质!贵州大学通过多组学揭秘超级稻最优施氮机制
2026 年 6 月 25 日
香蕉是支撑我国热区产业发展与全球粮食供给的核心作物,但稳产保收长期受香蕉枯萎病的严重制约。长期以来,生防菌体外抑菌效果亮眼,但是一到田间应用就效果不稳。根际微生物是植物抵御土传病害的天然防线,根系分泌的代谢物则是调控菌群功能的核心信号,直接决定生防体系的田间稳定性。链霉菌如何通过植物代谢重塑根际菌群实现抗病,绿色防控与稳产提质能否通过微生态调控实现平衡都是亟待解决的问题。
2026年5月,中国热带农业科学院谢江辉研究员、胡会刚研究员团队联合安徽农业大学王伟教授团队在Microbiome发表了一篇题为 “Streptomyces‑induced Glycerol‑3‑Phosphate enriches beneficial microbiota to enhance resistance against banana Fusarium wilt” 的研究论文。本研究结合根际微生物组与植物代谢组多组学联合分析,系统探究了海洋源马来西亚链霉菌WHL7调控香蕉根际微生态、增强枯萎病抗性的代谢特征,以阐明甘油-3-磷酸介导根际有益菌群富集的生理调控机制。深入理解生防微生物调控作物抗病的作用机制,将有助于优化香蕉枯萎病绿色防控策略,实现抗病与稳产协同提升。
百谱生物提供了非靶向代谢组检测与分析服务!

文章题目:Streptomyces-induced Glycerol-3-Phosphate enriches beneficial microbiota to enhance resistance against banana Fusarium wilt
期刊名称:Microbiome
影响因子:14.9
发表时间:2026年5月
发表单位:中国热带农业科学院、安徽农业大学
研究方法:基因组+微生物组+非靶向代谢组
研究思路:
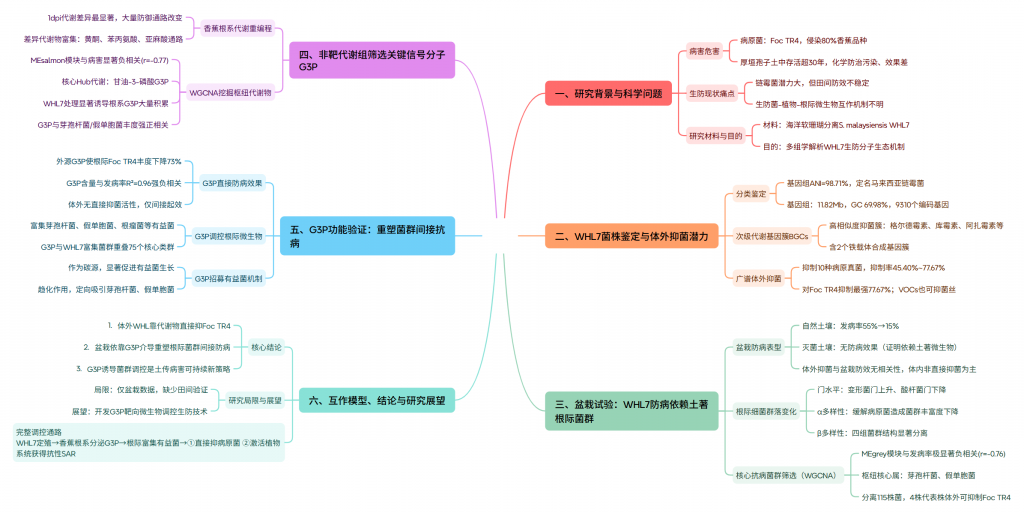
材料与方法
1. 实验材料
选选用香蕉主栽品种与核心功能菌株:
- 植物材料:巴西蕉组培苗
- 生防菌株:马来西亚链霉菌WHL7
- 病原菌:Foc TR4,含GFP荧光标记株
2. 实验设计
温室盆栽设置两组独立验证实验:
- WHL7生防实验:设空白对照、单接Foc TR4、单施WHL7、WHL7+Foc TR4 共4组,自然土壤与灭菌土壤平行开展,培养40天后取样
- G3P功能验证:设0、0.75、1.5、3.0 nM共4个G3P浓度梯度,蕉苗预处理2周后接种病原菌,14天后取样检测
3. 检测维度
研究从表型到分子层层递进解析:
- 菌株基因组与功能:全基因组测序、次生代谢基因簇预测、体外广谱抑菌活性测定
- 植株表型生理:发病率与病情指数、生长指标、病原菌侵染观察与丰度定量
- 根际微生物组:16S rRNA高通量测序、群落多样性分析、抗病核心菌群网络挖掘
- 根部代谢组:非靶向代谢组检测、差异代谢物鉴定、防御通路富集分析
- 互作机制验证:根际有益菌分离、G3P促生长与趋化实验
主要结果
一、基因组学:WHL7携带丰富抗真菌次生代谢基因簇
系统发育与ANI分析证实,WHL7为马来西亚链霉菌(Streptomyces malaysiensis),其基因组大小约11.82Mb,GC含量69.98%,共预测到9310个蛋白编码基因,其中61.45%的注释基因集中在代谢功能分类,符合链霉菌次生代谢活跃的典型特征。通过antiSMASH分析,基因组中鉴定出多个已证实抗真菌功能的次生代谢基因簇,包括格尔德霉素、库马霉素、尼日利亚菌素、阿扎霉素F3a等,相似度最高达100%,同时还存在2个铁载体合成基因簇,从基因组层面解释了其体外广谱抑制病原真菌的能力——对Foc TR4的体外菌丝生长抑制率可达77.67%。
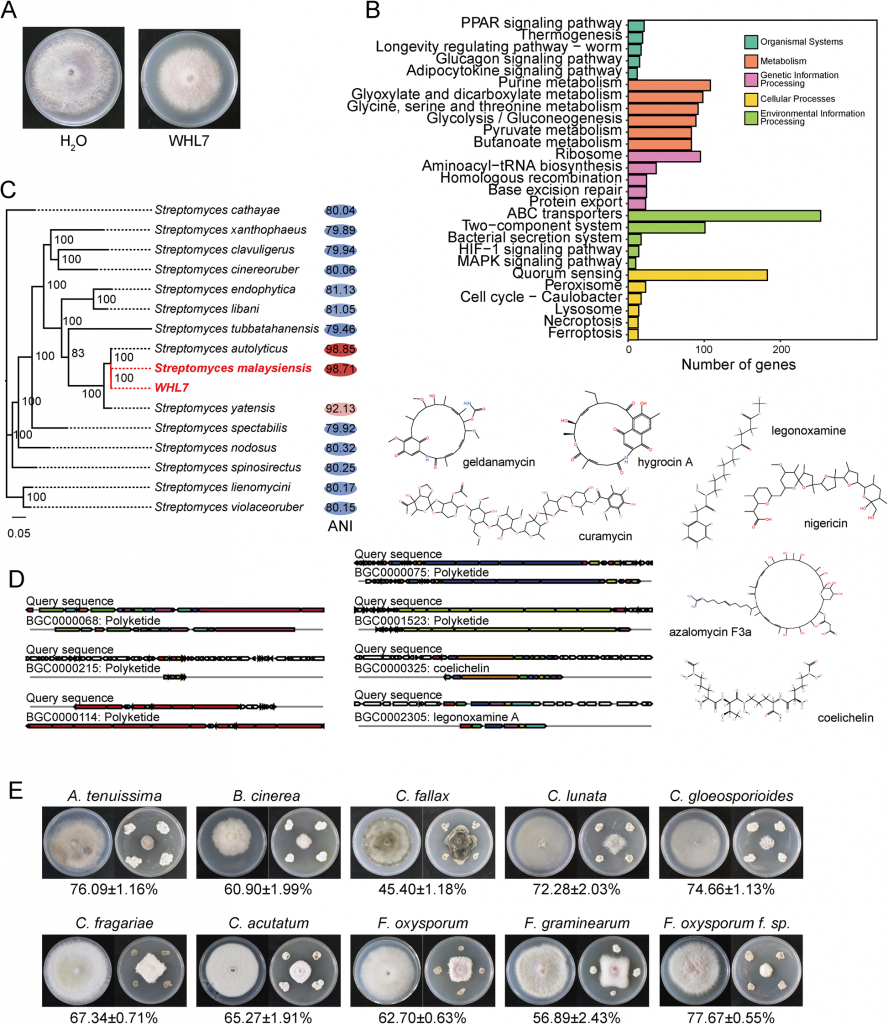
图1 WHL7的抗真菌活性及分类学鉴定
二、微生物组学:重塑根际菌群才是核心
盆栽实验发现了关键的机制线索:WHL7在自然土壤中能将枯萎病发病率从55%降至15%,但在灭菌土壤中完全失去保护效果,且WHL7自身在根际的丰度并未随处理显著提升。这直接说明,其生防作用并非依赖自身定殖直接抑菌,而是通过调控土著微生物组间接实现。
16S rRNA测序结果显示,Foc TR4侵染会导致根际微生物多样性显著下降,而WHL7处理能有效缓解这种多样性降低的趋势,且菌群结构发生明显重塑:变形菌门丰度显著上升、酸杆菌门丰度下降。结合差异丰度分析与WGCNA共表达网络,研究锁定了与抗病强相关的核心模块MEgrey,该模块与发病率呈显著负相关(r=-0.76, p=0.004),其中的核心枢纽菌属为芽孢杆菌(Bacillus)和假单胞菌(Pseudomonas)。后续分离培养验证也证实,这两类菌均对Foc TR4有显著的体外拮抗活性,是WHL7诱导的核心功能有益菌群。
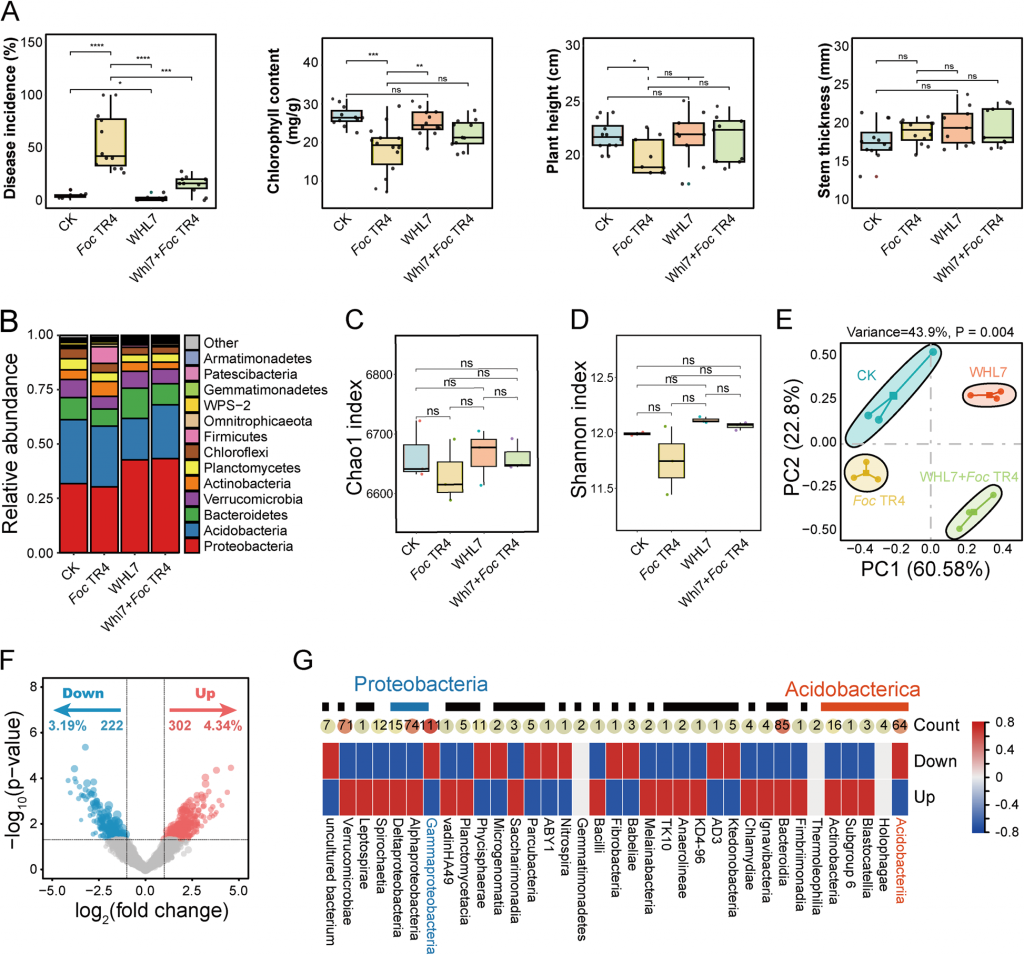
图2 WHL7处理后的生理特征及细菌群落结构
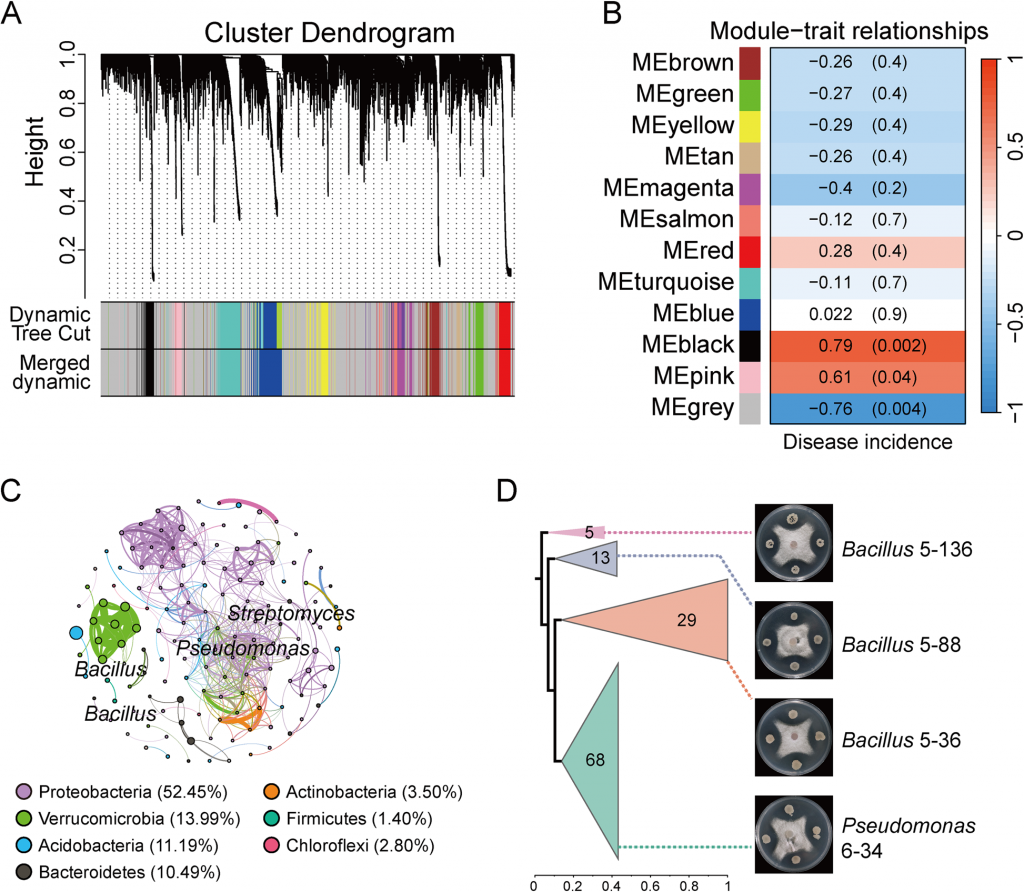
图3 通过WGCNA分析鉴定疾病相关的ASV
三、代谢组学:找到关键信号分子-G3P
为了明确WHL7引导根际菌群富集的分子媒介,研究团队对香蕉根部进行了非靶向代谢组检测。结果显示,WHL7处理会引发香蕉根部的代谢重编程,在病原菌侵染早期(1天)代谢差异最为显著,差异代谢物显著富集于黄酮类生物合成、苯丙氨酸/酪氨酸代谢、α-亚麻酸代谢等经典植物防御相关通路。
通过代谢组WGCNA分析,研究人员锁定了与发病率强负相关的MEsalmon模块(r=-0.77, p=0.003),并通过拓扑算法筛选出核心枢纽代谢物甘油-3-磷酸(G3P)。菌群-代谢物关联分析显示,G3P的丰度与芽孢杆菌、假单胞菌等多种有益菌的丰度呈显著正相关,且相关性强度高于其他代谢物;同时WHL7处理能显著诱导香蕉根部G3P的积累,说明 G3P很可能是植物传递给根际菌群的 “招募信号”。
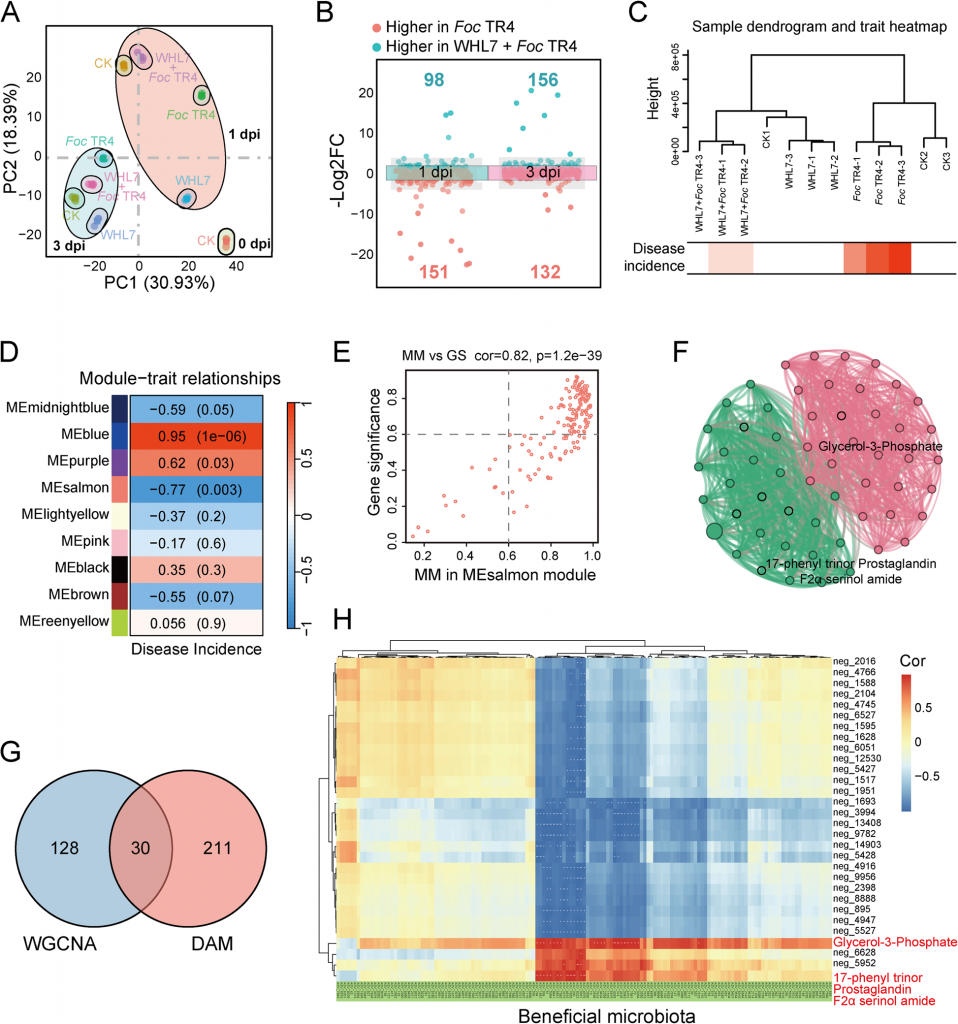
图4 香蕉根中差异积累代谢物分析
四、多组学联合验证:G3P定向招募有益菌,间接实现抗病
为了验证G3P的功能,研究人员开展了系统的外源施加实验,完整验证了 “G3P 富集有益菌→抑制病原菌→减轻病害” 的逻辑链:
体外实验证实,G3P本身无法直接抑制Foc TR4生长,说明其抗病作用是间接的;
盆栽实验显示,外源G3P处理能显著降低根际Foc TR4的丰度(最高降低73%),减少病原菌对球茎和根部的侵染,且病害发生率与G3P含量呈极强负相关(R²=0.96);
微生物组检测发现,外源G3P同样能定向富集芽孢杆菌、假单胞菌,且与WHL7处理富集的有益菌群重合度达52.45%;
趋化与生长实验进一步证实,G3P可作为易利用碳源促进芽孢杆菌、假单胞菌的增殖,同时诱导其趋化运动,直接解释了“菌群富集”的底层机制。
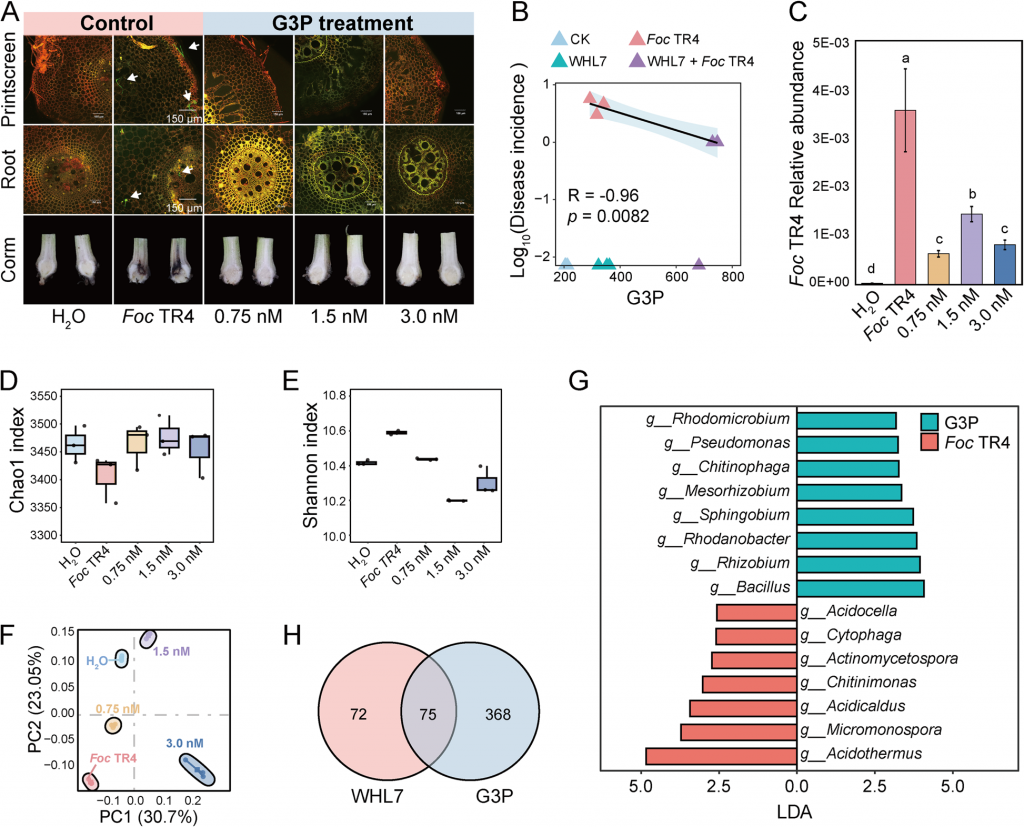
图5 G3P通过调节香蕉根际微生物群落抑制Foc TR4感染
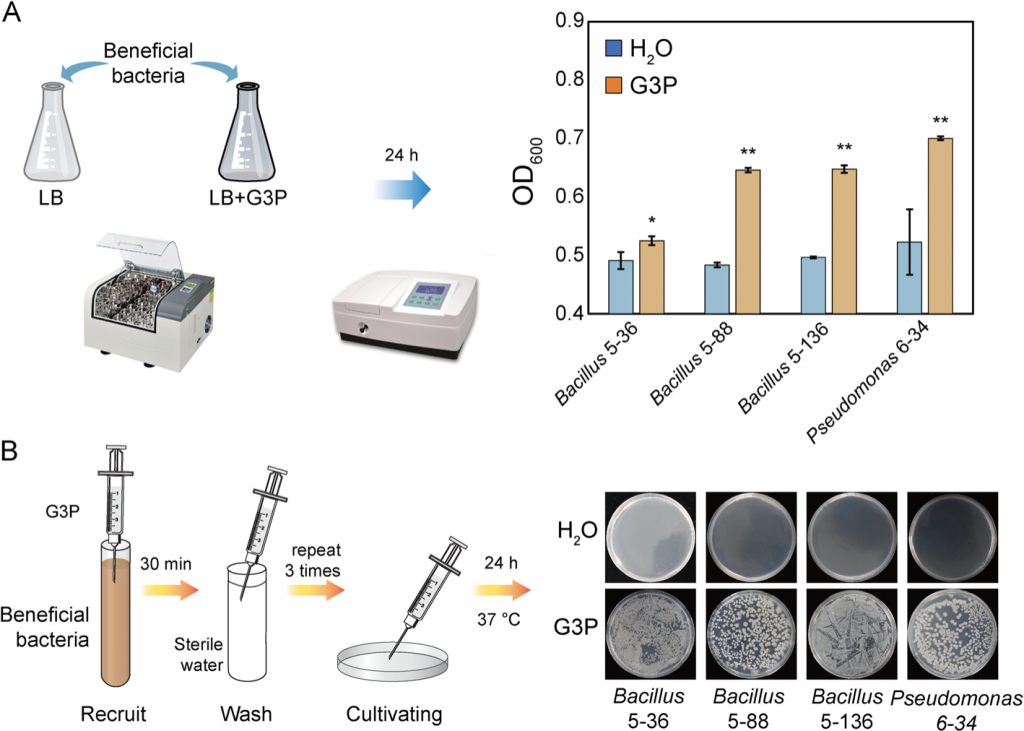
图6 通过G3P促进有益细菌的生长及趋化性
研究总结
本研究证实WHL7在体外具有广谱抗真菌活性,尤其对Foc TR4菌株效果显著。药效学实验表明,WHL7在自然土壤中的抗真菌机制与其平板试验中的拮抗作用模式存在差异。代谢组学分析显示,接种WHL7可刺激植物根系分泌G3P,从而富集根际有益微生物(尤其是芽孢杆菌属和假单胞菌属)。虽然G3P本身并不直接抑制Foc TR4的生长,但外源施用该物质能重现WHL7诱导的微生物群落变化并降低病原菌定殖水平。这些富集的微生物通过关键代谢途径激活植物的系统性获得性抗性。研究结果表明,WHL7并非通过直接拮抗作用,而是通过诱导G3P介导的有益细菌富集来间接抑制Foc TR4。
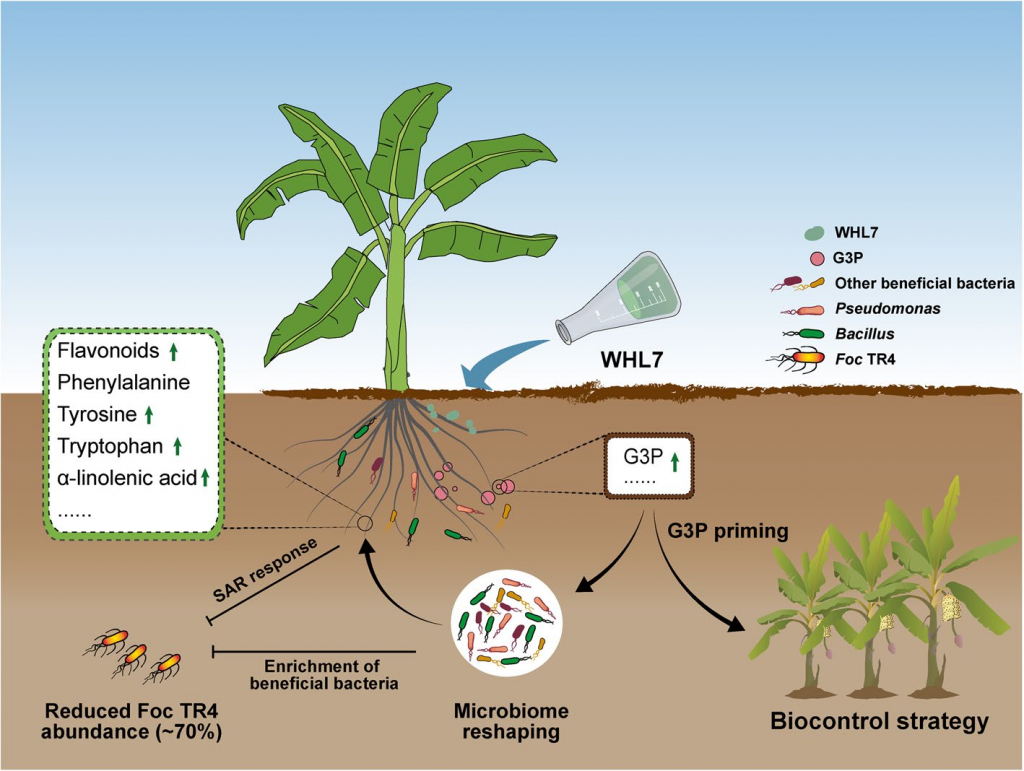
图7 WHL7植物-微生物组间的相互作用模型



{kind=link}
{kind=link}
{kind=link}